13. О возникновении и развитии жизни на Земле
На основании того, что было сказано в предыдущей главе, мы можем с достаточной для наших целей строгостью и точностью определить "живое вещество" как такой сложный молекулярный агрегат, в котором имеется "управляющая система", включающая в себя механизм передачи наследственной информации, обеспечивающей сохраняющие реакции следующим поколениям. Тем самым благодаря неизбежным "помехам" при передаче такой информации наш молекулярный комплекс ("организм") способен к мутациям, а следовательно, к эволюции.
Возникновению живого вещества на Земле (и, как можно судить по аналогии, на других планетах) предшествовала довольно длительная и сложная эволюция химического состава атмосферы, в конечном итоге приведшая к образованию органических молекул. Эти молекулы впоследствии послужили как бы "кирпичами" для образования живого вещества.
Коль скоро, согласно всем существующим космогоническим гипотезам, планеты образуются из первичной газово-пылевой субстанции, химический состав которой аналогичен химическому составу Солнца и звезд, первоначальная их атмосфера состояла в основном из простейших соединений водорода - наиболее обильного элемента в космосе. Больше всего было молекул Н2, Н2О, СО2, NH3 и СН4. Кроме того, первичная атмосфера должна была быть богата инертными газами, прежде всего гелием и неоном. Тот простой факт, что в настоящее время обилие благородных газов на Земле по сравнению с Солнцем ничтожно мало (В земной атмосфере имеется довольно значительное количество (около 1%) аргона. Однако атмосферный аргон образовался позже в результате радиоактивного распада калия и никакого отношения к первоначальной атмосфере не имеет)), означает, что они в свое время диссипировали в межпланетное пространство. Вместе с ними должны были диссипировать и молекулы, содержащие водород.
Для понимания эволюции планетных атмосфер особенное значение имеет анализ содержания благородных газов и их изотопов в атмосферах планет земной группы. Это следут из химической инертности этих газов в сочетании с тем, что тяготение планеты должно их удержать в атмосфере в течение всего времени эволюции атмосферы (за исключением легкого гелия). Выполненный советскими учеными во время полета "Венеры-11" и "Венеры-12" изотопный анализ атмосферы нашей космической соседки дает для этого богатый материал. В табл. 4 приведено относительное содержание разных изотопов благородных газов в атмосферах планет земной группы.
| - | 36А (см3/г) | 40А/36A | 40А (см3/г) | 36A/38A | 20Ne (см3/г) | 84Kr (см3/г) | 132Xe (см3/г) | 36A/84Kr |
| Венера | 2,2•10-6 | 1,2 | 2,6•10-6 | 5,0 | 5,3•10-7 | 2,6•10-8 | - | 120 |
| Земля | 2,1•10-8 | 296 | 6,2•10-6 | 5,0 | 1,1•10-8 | 4,3•10-10 | 1,6•10-11 | 49 |
| Марс | 1,0•10-10 | 3000 | 3,0•10-7 | - | 8,0•10-11 | 5.0•10-12 | 7,5•10-13 | 32 |
Обращает на себя внимание, что абсолютное содержание изотопа аргона 40А в атмосферах Земли и Венеры довольно близко. Так и должно быть, так как этот изотоп непрерывно образуется из изотопа калия, довольно обильного в коре обеих планет. Также понятно, почему в атмосфере Марса количество 40А на порядок меньше, чем в атмосферах Земли и Венеры - ведь масса Марса меньше. Совершенно неожиданно, однако, что "нерадиогенный" изотоп 36А в атмосфере Венеры так же обилен, как и радиогенный изотоп 40 А. Между тем в атмосферах Земли и Марса обилие изотопа 36А в сотни раз меньше, чем 40А. Столь разительное различие должно иметь глубокий космогонический смысл, т. е. оно должно отражать условия образования планет солнечной системы и их атмосфер. М. Н. Изаков из наблюдаемого изотопного состава атмосфер "внутренних" планет делает весьма радикальный вывод, что атмосфера Венеры была "захвачена" из протопланетного облака, между тем как на Земле и особенно на Марсе основная часть атмосферы имеет вторичное происхождение и обусловлена "дегазацией" пород, образующих кору этих планет. Этот важный вывод нуждается, однако, в подтверждении.
Необходимо сразу же подчеркнуть, что современная атмосфера нашей Земли совершенно уникальна. Сейчас уже благодаря выдающимся успехам космонавтики, мы надежно знаем состав атмосфер всех планет земной группы. Подробно об этом речь будет идти в гл. 16. Сейчас мы только подчеркнем, что основным газом в современных атмосферах Марса и Венеры является углекислота (свыше 95%). Между тем свободного кислорода в чрезвычайно разреженной атмосфере Марса всего лишь 0,2%, а на Венере и того меньше.
В земной атмосфере углекислота составляет совершенно ничтожную долю - 0,032% (К сожалению, этот процент растет благодаря неконтролируемому промышленному развитию)). В то же время вулканическая деятельность нашей планеты (так же как Венеры и Марса) щедро поставляет в атмосферу СО2. Куда же исчез углекислый газ? Почему в атмосферах наших "соседей" по Солнечной системе он постепенно накопился, а у нас "исчез"? Ответ на этот вопрос однозначен: это произошло благодаря жизнедеятельности покрывающих всю нашу планету растений, которые, используя солнечную энергию с помощью хлорофилла, из нескольких молекул СО2 и H2О синтезируют глюкозу. Освободившиеся молекулы кислорода при этом непрерывно поступают в атмосферу. Таким образом, в течение долгой истории Земли благодаря растениям (а также химическим реакциям на поверхности Земли) земная атмосфера была практически "очищена" от СО2 и насыщена молекулами О2.
Кислород в земной атмосфере находится в состоянии динамического равновесия. Если бы не жизнедеятельность растений (они поставляют в атмосферу ежегодно 1011 тонн кислорода), исключительно активные молекулы этого элемента вступили бы в различные химические реакции и исчезли бы из нашей атмосферы за какие-нибудь 10 000 лет! (Человечество варварски относится к сохранению этого чуда - насыщенной кислородом земной атмосферы. Сплошная вырубка лесов (особенно в Бразильской сельве), а также хаотическое промышленное развитие уже сейчас нарушили кислородный баланс нашей планеты. Можно, конечно, утешаться, что на несколько тысяч лет кислорода еще хватит. Однако существ, исповедывающих такую "философию" ("после нас - хоть потоп"), вряд ли следует причислять к виду "Homo Sapiens")).
С точки зрения планетолога современная атмосфера Земли представляет собой "астрономический нонсенс" или, проще говоря, чудо. Это надо же - 21 % атмосферы состоит из немыслимо химически активного газа. И все это - результат развития жизни на нашей планете! Этот пример со всей наглядностью показывает как развитие жизни на планете приводит к космическим последствиям. В дальнейшем, в частности, при анализе проблем разумной жизни во Вселенной, мы будем возвращаться к этому вопросу неоднократно.
Сколько же времени на Земле существовала первичная атмосфера? Имеются довольно надежные геологические и геохимические данные, указывающие на то, что уже 3,5 млрд. лет назад земная атмосфера была довольно богата кислородом. Жизнь должна была возникнуть на Земле задолго до того, как атмосфера стала богата кислородом, так как последний является продуктом жизнедеятельности растений. Согласно недавней оценке Сагана, жизнь на Земле возникла 4,2±0,2 млрд. лет назад.
Эта оценка следует из того, что самые древние из известных на Земле организмов - сине-зеленые водоросли имеют возраст 3,2 млрд. лет. Так как эти организмы довольно сложны, ясно, что от момента зарождения жизни на Земле до их возникновения прошло немало времени. Другими словами, уже на ранних фазах эволюции Земли на ней возникала жизнь.
Схематически путь эволюции органического вещества на Земле можно представить в виде следующей таблицы:
| I Образование Земли | II Возникновение живых систем. Клетка | III Эволюция одноклеточных. Возникновение клеточной дифференциации | IV Эволюция многоклеточных | V Человек |
Рассмотрим теперь более подробно начальные этапы этой эволюции. По-видимому, наибольшие загадки ставит перед нами переход от первой фазы эволюции ко второй.
В настоящее время накоплен значительный экспериментальный материал, иллюстрирующий, каким образом такие простые вещества, как вода, метан, аммиак, окись углерода, аммонийные и фосфатные соединения и др., превращаются в высокоорганизованные структуры, являющиеся основными строительными блоками клетки - единицы живого. Эти опыты, начатые впервые американскими учеными Кельвином, Миллером и Юри, положили начало новому научному направлению, получившему впоследствии название "пребиологической химии".
Так, например, опытами Миллера доказано, что при прохождении электрических разрядов через смесь метана (СН4), молекулярного водорода (Н2), аммиака (NH3) и паров воды (эта смесь довольно хорошо моделирует первичную атмосферу Земли) возникали глицин, аланин и другие аминокислоты, а также ряд органических соединений. Точно так же экспериментально доказано, что в такой смеси образование органических соединений (в частности, аминокислот) может происходить под воздействием ультрафиолетовой радиации. Можно полагать, что в условиях неокисленной земной атмосферы, когда ультрафиолетовое излучение Солнца беспрепятственно могло достигать земной поверхности (В настоящее время близкая ультрафиолетовая часть солнечного спектра поглощается озоном О3, а более далекая - молекулами кислорода О2 и азота N2)), важная роль в образовании первых органических соединений принадлежала этому источнику энергии. В то же время серьезное значение могли иметь и другие источники энергии. Следует подчеркнуть, что уже первые попытки экспериментального изучения проблемы возникновения жизни на молекулярном уровне продемонстрировали возможность многочисленных "вариантов", которые могли иметь место в течение первого миллиарда лет истории Земли.
Таким образом, можно считать доказанным, что под воздействием различных форм энергии на примитивной Земле возникали достаточно сложно организованные органические молекулы.
В синтезе органики наиболее существенную роль должны были играть электрические разряды, ударные волны, ультрафиолетовое излучение Солнца, вулканическое тепло, радиоактивный распад 40К.
Из табл. видно, что основной вклад в процессы абиогенного синтеза вносит ультрафиолетовое излучение Солнца. Однако вопрос об относительной эффективности различных видов энергии не так прост, как это кажется на первый взгляд. В экспериментах по абиогенному синтезу были использованы все источники энергии, перечисленные в табл. 6. При этом выяснилось, что определяющим моментом является не общее количество энергии, а "к. п. д." той или иной модели образования органических веществ.
| Источник | Средняя энергия на всю поверхность Земди (в единицах 1020 кал/год) |
| Распад 40K (в настоящее время) | 0,3 |
| Распад 40K (2,6•109 лет назад) | 1,2 |
| Ультрафиолетовое излучение Солнца (λ<1500Å) | 0,08 |
| Ультрафиолетовое излучение Солнца (λ<2000Å) | 4,5 |
| Вулканизм (лава 1000°С) | 0,04 |
| Удары метеоритов | 0,05 |
| Молнии | 0,05 |
Полезно рассмотреть следующие этапы в эволюции органического вещества на примитивной Земле:
1. Эволюция малых молекул.
2. Образование полимеров.
3. Возникновение каталитических функций.
4. Самосборка молекул.
5. Возникновение мембран и доклеточная организация.
6. Возникновение механизма наследственности.
7. Возникновение клетки.
Необходимо отметить, что в настоящее время не представляется возможным искусственно воспроизвести в лабораторных условиях возникновение механизма матричного копирования, реализуемого в живой клетке нуклеиновыми кислотами. Между тем, по-видимому, в этом состоит суть проблемы возникновения жизни на Земле.
Наиболее изученным этапом в пребиологической химии является эволюция малых молекул. Было экспериментально изучено воздействие всех вышеперечисленных видов энергии на смеси различных газов: водород, метан, аммиак, окись углерода, двуокись углерода, азот, вода, кислород, сероводород. При этом было установлено, что если смесь не была окислительной, то всегда образовывались аминокислоты и другие биологически активные соединения.
Определяющими промежуточными продуктами в синтезе аминокислот, оснований нуклеиновых кислот, Сахаров и порфиринов являются формальдегид и цианистый водород. Образование этих простых продуктов происходит и в газовой, и в водной фазе. Образование же более сложных молекул (аминокислот) происходит главным образом в водной среде.
Среди возможных механизмов образования аминокислот можно указать на синтез Штрекера, как конечный этап превращения аминонитрилов и циангидринов
NH3+KCHO+HCN⇔NH2CH(K)CN+H2O (аминонитрил),
KCH(NH2)CN-f 2Н2O->KCH(NH2)COOH+NH3 (циангидрин).
Что касается синтеза оснований нуклеиновых кислот, то здесь также, как выяснилось, центральную роль играет цианистый водород. Так, при синтезе аденина "суммарную" реакцию образования этого соединения можно записать следующим образом:
5HCN -> аденин.
При образовании Сахаров в условиях, моделирующих примитивную Землю, происходит щелочная конденсация формальдегида. Протекание этой реакции катализируется гидроокисями щелочноземельных металлов.
В экспериментах, проведенных Гейбелом и Поннамперумой, водные растворы формальдегида в различных концентрациях нагревались в присутствии каолинита, который использовался в качестве природного катализатора. В числе продуктов реакции были отождествлены триозы, тетрозы, пентозы, гексозы. Была отождествлена также рибоза.
Чрезвычайно важной группой соединений, присутствующих в большинстве живых организмов, являются порфирины. Порфириновая структура лежит в основе хлорофилла. Целый ряд важнейших ферментов, таких как каталаза, пероксидаза и др., также имеют порфириновую структуру.
В экспериментах по абиогенному синтезу порфирин был идентифицирован как один из продуктов реакции в смеси метан - аммиак - вода - водород под действием электрического разряда.
Наиболее существенным достижением в области пребиологической химии можно считать абиогенный синтез нуклеотидов и полинуклеотидов, осуществленный впервые Шраммом из углеводов и гетероциклических оснований с помощью метафосфорных эфиров (МФЭ). В процессе синтеза образовывались продукты различной молекулярной массы и структуры, причем нуклеотиды в полинук-леотидной цепи располагались случайно, не образуя какой-либо определенной последовательности.
Значительный интерес представляют также эксперименты Фокса по термической полимеризации аминокислот.
Таким образом, многочисленные эксперименты по абиогенному синтезу продемонстрировали возможность образования основных классов биологических активных соединений небиологическим путем в условиях, моделирующих природные условия, существовавшие на примитивной Земле.
Однако образование самых сложных молекул не решает вопроса об отборе и сохранении определенных типов молекулярных соединений. На определенной стадии усложнения структуры молекул возникает такое принципиально новое свойство их как возвратный катализ. Образовавшиеся довольно сложные молекулы должны разрушаться (диссоциировать) при поглощении более длинноволнового излучения, чем то, которое стимулировало их образование. Так как поток солнечного излучения в области более длинных волн значительно превосходит поток ультрафиолетового излучения, стимулирующего синтез первичных органических соединений, последние будут разрушаться, и какого-либо накопления их происходить не должно. Заметим, что эта трудность является общей для всех механизмов образования первичных органических соединений, так как неокисленная атмосфера планеты должна быть прозрачной для ультрафиолетовых лучей Солнца. Сейчас намечается несколько путей преодоления этой трудности. Например, можно предположить, что после сформирования гидросферы образовавшиеся в ее поверхностных слоях органические соединения путем конвекции переносились на достаточную глубину, куда уже "разрушительное" излучение не доходило.
Зная поток ультрафиолетового излучения Солнца, стимулирующего образование органических веществ, и считая, что вновь образовавшиеся вещества не разрушаются, а постепенно накапливаются, можно оценить количество образующегося таким способом органического вещества на Земле. Такие вычисления произвел Саган, который в предположении, что этот процесс длился 1 млрд. лет, нашел, что над каждым квадратным сантиметром земной поверхности могло образоваться несколько килограммов органических соединений. Эта величина представляется достаточно большой. Например, если бы все эти образовавшиеся в раннюю эпоху развития нашей планеты органические вещества растворить в мировом океане, концентрация такого раствора была бы приблизительно 1%.
Так как есть основания полагать, что объем мирового океана за геологическую историю Земли почти не менялся, можно сделать вывод, что первобытный океан представлял собой 1%-ный раствор различных органических соединений. Довольно крепкий питательный бульон! Эта среда была весьма благоприятна для образования новых, более сложных органических соединений. В частности, из аминокислот могли синтезироваться различные белковые соединения.
До сих пор предполагалось, что жизнь как-то возникла на всей "осредненной" поверхности первобытной Земли, для чего потребовались сотни миллионов лет. Но, конечно, это могло быть и наверняка было не так. В отдельных местах земной поверхности условия для эволюции сложных молекул в первые примитивные формы жизни могли быть особенно благоприятны. Идеи "локального" возникновения жизни на Земле и притом в сравнительно короткие сроки высказывались неоднократно. Недавно Л. М. Мухин предложил интересную гипотезу, что жизнь могла возникнуть в области подводных вулканов.
По всей видимости, именно подводный вулканизм мог играть известную роль в образовании предшественников сложных органических молекул. Действующий вулкан можно рассматривать не только как источник тепла, но и как источник простых соединений, таких как СО, СН4, NH3, CО2, H2О, H2, H2S и пр., необходимых для синтеза органического вещества. Реакции, происходящие между этими газами в условиях повышенных температур и давлений, должны приводить к образованию предшественников сложных органических соединений, цианистого водорода и формальдегида. Гидросфера (океан) используется в данной модели как фактор, обеспечивающий стабильность образовавшихся продуктов вследствие больших перепадов температуры в зоне действия подводного вулкана. Кроме того, в области подводного вулкана имеется широкий диапазон давлений, что весьма существенно, так как высокие давления необходимы для повышения выхода продукта в ряде реакций. Наконец, наличие в области подводного извержения зон с температурой 50-100°С обеспечивает прохождение ряда реакций, приводящих к синтезу более сложных органических соединений. Механизмы этих реакций освещены в работах Оро и Поннамперумы.
Образование в процессе извержения твердых частиц обусловливает наличие катализаторов и может способствовать в дальнейшем процессам концентрирования и полимеризации органики.
Л. М. Мухин указывает на некоторые реакции, которые могут иметь место в зоне подводных извержений:
катализаторы
1) CH4+NH3->HCN+3H2,
катализаторы
2) 2CO+NH3->HCN+CО2+H2,
катализаторы
3) CO+H2->альдегиды и другие кислородосодержащие соединения, углеводороды.
Таким образом, вследствие возможного образования в зоне действия подводного вулкана HCN и СН2О, подводные вулканические процессы можно рассматривать как источник небиологического синтеза сложных органических соединений.
Рассмотрим теперь некоторые численные значения, которые носят характер ориентировочных оценок. Масса газа, выброшенного при сильном извержении, имеет порядок величины 1012 г. Если принять, что в течение истории развития Земли такие извержения были ежегодно, то при благоприятных условиях могло образоваться до 1017 г органических соединений.
"Вулканический" механизм образования сложных молекул может иметь принципиальное значение в условиях, где по ряду причин воздействием ультрафиолетового излучения на исходные материалы можно пренебречь.
Предложенный Мухиным механизм образования сложных молекул не требует наличия метано-аммиачной атмосферы. Было бы интересно проверить этот механизм экспериментально в зоне действия какого-либо подводного вулкана.
Много лет тому назад Бернал высказывался в том смысле, что жизнь могла зародиться в иле небольших лагун. В таких условиях полимеризация молекул может протекать гораздо быстрее, так как микроскопические частицы ила могут выступать в роли своеобразных катализаторов. Это предположение Бернал а было подтверждено экспериментально. Любопытно отметить, что сложные органические молекулы значительно лучше "сопротивляются" разрушительному воздействию ультрафиолетовых лучей, а также нагреву, чем простые. Поэтому следует ожидать, что с течением времени должны "выживать" более сложные молекулы, в то время как простые должны разрушаться. Довольно любопытный пример "естественного отбора" у неживой материи!...
Наряду с описанным "естественным отбором", приводящим к преимущественному образованию сложных органических соединений, будут происходить, и притом довольно эффективно, "сливания" таких молекул в целые молекулярные агрегаты, насчитывающие сотни тысяч и миллионы молекул. Такие образования называются "коацерватными каплями". Они неоднократно исследовались экспериментально. На рис. 38 приведена фотография таких капель, сделанная через микроскоп при увеличении в 320 раз. В итоге образования коацерватных капель в них могут быть сконцентрированы все белковые молекулы, присутствующие в мировом океане - растворе. В окружающей воде будут растворены только сравнительно простые, низкомолекулярные соединения.
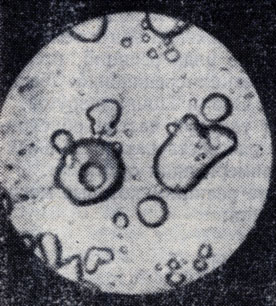
Рис. 38. Коацерватные капли (увеличено в 320 раз)
Академик А. И. Опарин считает, что именно эти коацерватные капли при определенных условиях могли дать начало образованию первичных живых систем. Об этом свидетельствует ряд интересных свойств коацерватных капель, ставших известными в результате лабораторных исследований. В частности, эти капли обладают свойством улавливать и впитывать в свою структуру некоторые вещества из окружающего их низкомолекулярного раствора. В этом А. И. Опарин усматривает зачаточные формы процесса обмена веществ - важнейшего, по его мнению, атрибута жизни. Он подчеркивает, что в мире коацерватов имеют место полные аналоги процесса естественного отбора. По этому поводу он пишет: "Образовавшиеся в земной гидросфере коацерватные капли находились погруженными не просто в воде, а в растворе разнообразных органических веществ и неорганических солей. Эти вещества и соли адсорбировались коацерватными каплями и затем вступали в химическое взаимодействие с веществом самого коацервата. Происходили процессы синтеза. Но параллельно с ними шли и процессы распада. Скорость как тех, так и других процессов зависела от внутренней организации каждой данной капли. Более или менее длительно существовать могли только капли, обладавшие известной динамической устойчивостью, в которых при данных условиях внешней среды скорости синтетических процессов преобладали над скоростями разложения. В обратном случае капли были обречены на исчезновение. Индивидуальная история таких капель быстро обрывалась, и поэтому такие "плохо организованные капли" уже не играли никакой роли в ходе дальнейшей эволюции органической материи" (А. И. Опарин, В. Г. Фесенков. Жизнь во Вселенной. - М.: Изд-во АН СССР, 1956)).
С гипотезой А. И. Опарина в настоящее время трудно согласиться. Наличие аналогов обмена веществ и "естественного отбора" у коацерватов еще не есть доказательство того, что они могли привести к образованию первых примитивных живых организмов. Основными свойствами всякого живого организма, помимо обмена веществ, являются наличие "копировальной системы", "кода", передающего по наследству все характерные признаки данной особи. Между тем у коацерватов ничего подобного нет. Изобилие на первобытной земле всевозможных, в том числе и достаточно сложных, "строительных блоков", из которых построено все живое, еще не объясняет, как возникла и стала функционировать живая субстанция, представляющая собой даже в самых простых формах весьма сложную машину, а если говорить точнее, великолепно работающую современнейшую фабрику-автомат.
"Управляющая система" этой фабрики сосредоточена в одномерной структуре ДНК, хранящей информацию, записанную на языке, состоящем из четырех букв (оснований). Система осуществляет перевод этого языка на язык строящихся по ее командам белков, состоящий из 20 букв (аминокислот).
Как произошел качественный скачок от неживого к живому, гипотеза А. И. Опарина совершенно не объясняет. Только привлечение основных представлений современной молекулярной биологии, а также кибернетики, может помочь решению этой важнейшей, основной проблемы. Впрочем, пока не ясно, есть ли такое решение вообще.
Итак, центральной проблемой происхождения жизни на Земле является реконструкция эволюции механизма наследственности. Жизнь возникла только тогда, когда начал действовать механизм репликации. Ведь любая сколь угодно сложная комбинация аминокислот и других сложных органических соединений - это еще не живой организм. Можно, конечно, предположить, что при каких-то исключительно благоприятных обстоятельствах где-то на Земле возникла некая "праДНК", которая и послужила началом всему живому на Земле. Вряд ли, однако, это так, если гипотетическая "праДНК"былавполнеподобнасовременной.Деловтом, что современная ДНК сама по себе совершенно беспомощна. Она может функционировать только при наличии белков-ферментов. Думать, что чисто случайно, путем "перетряхивания" отдельных блоков - многоатомных молекул, могла возникнуть такая сложнейшая машина, как "праДНК" и нужный для ее функционирования комплекс белков-ферментов,- это значит верить в чудеса. Куда, например, более вероятно предположить, что какая-нибудь мартышка, беспорядочно барабаня по клавиатуре пишущей машинки, случайно напечатает 66-й сонет Шекспира... Выход из этого затруднительного положения может состоять в том, что сам репликационный механизм за первые сотни миллионов лет развития "пражизни" претерпел огромную эволюцию от простого к сложному. К сожалению, успехи в этой важнейшей области пока незначительны.
Рич, однако, указал на значительное сходство строения молекул ДНК и РНК, которые тем не менее выполняют в клетке совершенно различные функции. ДНК является носителем генетической информации, РНК служит для превращения этой информации в реальные молекулы белка, т. е. для непосредственного синтеза видовоспецифического белка.
Особого внимания заслуживает открытие у вируса табачной мозаики и у некоторых других вирусов не двух, а только одной нуклеиновой кислоты, более простой - РНК. Эта РНК оказалась способной осуществлять функции обеих нуклеиновых кислот - передачи наследственной информации и синтеза белка.
Можно допустить, что обе нуклеиновые кислоты произошли от одной общей более примитивной молекулы. Усложняясь и специализируясь в процессе эволюции, эта "прануклеиновая" кислота превратилась в функционально различные типы молекул ДНК и РНК. Возможно, что этой первичной нуклеиновой кислотой могла быть молекула, близкая к более простой РНК. Подобно РНК вируса табачной мозаики она обладала способностью к передаче наследственной информации и к синтезу белка. Возможно также, что вирусы, содержащие только одну РНК (филогенетически более раннее образование), следует рассматривать как современные модификации древней, примитивной формы жизни.
Все это может пролить некоторый свет на пути возникновения и развития живых существ от более простых форм управления и примитивной жизни к более сложным формам. Если небелковая ("неживая") молекула РНК в подходящей среде образует живые системы, то не на этом ли пути можно обнаружить "мостик" между неживой и живой природой? Решающее слово в этом важнейшем вопросе принадлежит будущим биохимическим и генетическим исследованиям.
Для образовавшихся на планете первых примитивных живых организмов высокие дозы жесткой радиации могут представлять смертельную опасность, так как мутации будут происходить так быстро, что естественный отбор не поспеет за ними.
Мы уже упоминали в гл. 5, что примерно один раз в сотни миллионов лет около Солнца вспыхивает сверхновая звезда, и в нашей планетной системе уровень космических лучей увеличивается в десятки и сотни раз. Однако для сравнительно короткоживущих примитивных жизненных форм такое увеличение уровня жесткой радиации не представляет серьезной опасности. Кроме того, длительность периодов повышенной интенсивности космических лучей сравнительно невелика (десятки тысяч лет). Другим возможным источником губительной жесткой радиации мог быть повышенный уровень радиоактивности на первобытной Земле. Однако расчеты показывают, что этот уровень вряд ли превышал современный более чем в 10 раз. Солнечное рентгеновское излучение в те времена, так же как и сейчас, не проникало через толщу атмосферы. И только один вид жесткой радиации имел высокую интенсивность - ультрафиолетовое излучение Солнца в области длин волн 0,29-0,24 мкм для которого первобытная атмосфера Земли, в отличие от современной, была прозрачной.
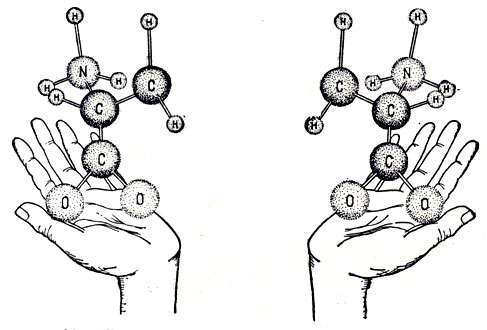
Рис. 39. 'Левые' и 'правые' органические молекулы
Так как Солнце в те времена излучало примерно так же, как и сейчас, мы можем оценить поток его излучения на Земле в указанной спектральной области. Этот поток оказывается равным 5•103 эрг/см2•с, т. е. примерно в 300 раз меньше полного потока солнечного излучения. Смертельная доза такой радиации для большинства современных микроорганизмов составляет 105-106 эрг/см2. Радиационная опасность отсутствует в том случае, когда за время жизни одного поколения живых организмов доза радиации меньше приведенной величины. Имеются некоторые основания полагать, что время жизни первобытных примитивных организмов было достаточно велико, например, несколько недель. Если считать, что для них доза в 103 эрг/см2 была опасной, то поток ультрафиолетовой радиации должен быть не больше 10-3 эрг/см2•сек, т. е. в 5 млн. раз меньше реального потока солнечного излучения. Отсюда следует важный вывод, что первичные живые организмы могли образоваться и развиваться только на достаточно большой глубине под водой. Слой воды в несколько десятков метров уменьшает поток ультрафиолетового излучения в десятки миллионов раз и тем самым обеспечивает необходимую для развития живых организмов "броню". Это является еще одним важным аргументом в пользу утверждения, что жизнь на нашей планете возникла и развивалась первоначально в воде, причем на достаточно большой глубине.
Мы остановились только на некоторых основных вопросах возникновения жизни на Земле и по аналогии - на других планетах. В этой проблеме еще очень много неясного. Например, все белковые соединения, входящие в состав живого вещества, имеют "левую асимметрию". Что это означает? Дело в том, что большое количество органических соединений может существовать в двух формах. Эти формы отличаются одна от другой противоположной ориентацией отдельных группировок атомов - некоторая группировка атомов в одной форме является зеркальным изображением соответствующей группировки в другой.
Когда происходит лабораторный синтез такого соединения, всегда "правые" и "левые" формы присутствуют в одинаковом количестве, так как "наращивание" молекул путем присоединения атомов и атомных группировок происходит случайным образом. Почему же в "живых" органических соединениях всегда присутствуют только "левые" формы?
Еще Пастер указал, что "асимметричный синтез" может происходить при наличии какого-нибудь природного асимметричного фактора. И действительно, если в лабораторных условиях синтезировать некоторые органические соединения под воздействием поляризованного по кругу света, то в зависимости от направления вращения светового вектора получаются преимущественно "правые" или "левые" формы синтезируемых веществ. К сожалению, таким способом трудно объяснить асимметрию "живых" молекул, так как в солнечном излучении отсутствует сколько-нибудь значительная составляющая, поляризованная по кругу. Впрочем, нельзя исключать того, что после прохождения значительной толщи первобытного океана, вода которого, быть может, обладала соответствующими оптическими свойствами, такая составляющая и возникала. Этот вопрос требует специального исследования.
Другой возможный путь асимметричного синтеза был указан Берналом. При синтезе некоторых органических веществ на поверхности оптически активных кристаллов (например, кварца) могут возникать формы определенной симметрии. Следует, однако, отметить, что в природе распространены как "правые", так и "левые" кристаллы. Поэтому ке совсем ясно, каким образом в живом веществе молекулы имеют асимметрию только одного знака и вряд ли асимметричный синтез в естественных условиях первобытной Земли мог происходить таким способом. Так или иначе, вопрос о причине асимметрии живой субстанции пока остается открытым.
Заслуживает внимания еще такой вопрос: почему жизнь на Земле не возникает из неживого вещества в настоящее время? И вообще - жизнь на Земле возникла однократно или многократно? Против возможностей повторного зарождения жизни на нашей планете из неживой субстанции можно выдвинуть такой серьезный аргумент: ранее возникшая жизнь не даст возможность новому зарождению жизни. Микроорганизмы и вирусы буквально съедят уже первые ростки новой жизни. Другим аргументом против "повторного" зарождения жизни является ничтожно малая вероятность этого процесса. Ведь нельзя исключить возможность того, что жизнь на Земле возникла случайно (см. ниже).
Существует еще одно обстоятельство, на которое, может быть, стоит обратить внимание. Хорошо известно, что все "живые" белки состоят из 20 аминокислот, между тем как всего аминокислот известно свыше 100. Не совсем понятно, чем отличаются эти 20 аминокислот от остальных своих "собратьев" (Впрочем, некоторые количества других аминокислот имеются у низших организмов. Следует, однако, заметить, что у этих организмов ДНК отличаются от обычных)). Нет ли какой-то глубокой связи между происхождением жизни и этим удивительным явлением? Мы еще раз должны подчеркнуть, что центральная проблема возникновения жизни на Земле - объяснение качественного скачка от "неживого" к "живому" - все еще далеко от ясности. Недаром один из основоположников современной молекулярной биологии проф. Крик на Бюраканском симпозиуме в сентябре 1971 г. сказал: "Мы не видим пути от первичного бульона до естественного отбора. Можно прийти к выводу, что происхождение жизни - чудо, но это свидетельствует только о нашем незнании".
Все же не будем отчаиваться - и эта твердыня непознанного будет взята; порукой этому является гигантский прогресс современной молекулярной биологии.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://12apr.su/ 'Библиотека по астрономии и космонавтике'